Archives
- 2026-08
- 2026-07
- 2026-06
- 2026-05
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-07
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-07
-
br Ultrasonic pretreatment of substrates Substrate pretreatm
2020-10-06
Ultrasonic pretreatment of substrates Substrate pretreatment is widely used in the biofuel, textile and food industries since the substrates are always difficult to degrade. During some enzymatic hydrolysis reactions, the protective layer of the substrate impedes the reaction. Ultrasonic treatmen
-
br Results On the th day the horizontal
2020-10-03
Results On the 8th day, the horizontal (Fig. 1) and vertical locomotor activity (Fig. 2) and the dorsal (Fig. 3) and indy 6 striatal dopamine release (Fig. 4) increased significantly in nicotine-treated rats, compared with the saline-treated rats. Tukey post-hoc test revealed the following p val
-
ionomycin receptor CPH models reported usually as lists of r
2020-10-02
CPH models, reported usually as lists of risk factors along with their parameters, are prevalent in medical literature. One such model is the CPH model created for the purpose of predicting the probability of one year survival of patients suffering from Pulmonary Arterial Hypertension [5]. The model
-
99026 br Agonist binding ETA activation is promoted
2020-10-01
Agonist binding ETA activation is promoted by binding of the endogenous peptidergic agonists to their orthosteric binding site on the receptor. ET1 and ET2 (Trp6-Leu7-ET1) bind with equal high affinity to ETA, whereas the third endogenous isopeptide ET3 (Thr2-Phe4-Thr5-Tyr6-Lys7-Tyr14-ET1) binds
-
Another interesting finding regarding substrate selectivity
2020-09-30
Another interesting finding regarding substrate selectivity of iPLA2-VIA in whole cellular systems stems from the observation that some of the major species hydrolyzed by the enzyme contain a 16:1 fatty Pregnenolone Carbonitrile sale at the sn-2 position [57,119], raising the possibility that iPLA2-
-
br Multi subunit RINGs There are RING
2020-09-30

Multi-subunit RINGs There are RING-type E3s that exist as multi-subunit assemblies (see Fig. 3B). A striking example is the Cullin RING Ligase (CRL) superfamily [35], which exhibits enormous plasticity in substrate specificity. Each CRL subfamily is characterized by a cullin protein (Cul-1, 2, 3,
-
BRCC is the catalytic subunit responsible for the majority
2020-09-30
BRCC36 is the catalytic subunit responsible for the majority of K63-Ub-specific DUB activity in the cytoplasm and in the nucleus as part of two distinct macromolecular assemblies characterized by the presence of either of the MPN– pseudo DUB proteins KIAA0157 or Abraxas (Cooper et al., 2009, Dong et
-
br Materials and methods br Results
2020-09-30
Materials and methods Results Discussion Regardless of obvious beneficial effects of GC on acceleration of foetal lung and cardiomyocytes maturation (Kamath-Rayne et al., 2012; Rog-Zielinska et al., 2015), the current study revealed that GC exposure during late pregnancy induces IR and alte
-
Fluorescein TSA Fluorescence System Kit australia Transducti
2020-09-30
Transduction of cellular signals by G protein-coupled receptors (GPCRs) is stringently regulated to prevent the deleterious effects of unrestrained GPCR signaling. The rapid termination of signaling mediated by agonist-occupied GPCRs is referred to as homologous desensitization and involves the foll
-
Regarding their reported beneficial properties
2020-09-29
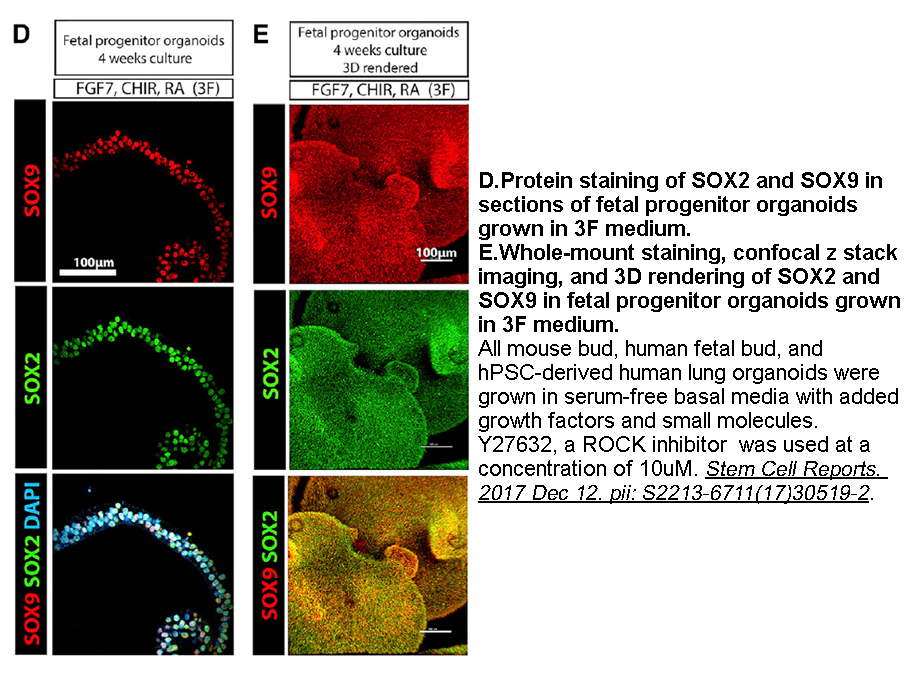
Regarding their reported beneficial properties on the digestive tract, the aim of this study was to evaluate the effect of water extracts of three Hypericum sps., HP, HA, HU and some of the main phenolic compounds (Ch, R and Q) on DNA protection against oxidative and alkylating DNA damage induced by
-
As shown in Fig there are two mechanisms for the
2020-09-29

As shown in Fig. 9, there are two mechanisms for the removal of the Va-acyl group from PC to make it available for incorporation into TAG with DGATs\' acting at the final acylation step. ①: Transfer of Va from PC to the acyl-CoA pool. This process can be driven by the reverse action of acyl- CoA:lys
-
The effect of NAT deletion on mitochondrial function in MDA
2020-09-29
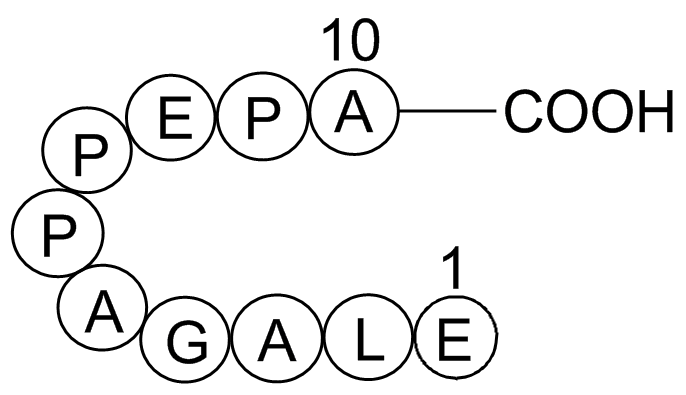
The effect of NAT1 deletion on mitochondrial function in MDA-MB-231 Pioglitazone has been reported elsewhere (Carlisle et al., 2018). However, unlike the data presented here, increases in reserve capacity and glycolytic reserve were seen. The reasons for this variance between our study and that of
-
br Concluding Remarks Synthetic cytokine biology has become
2020-09-29
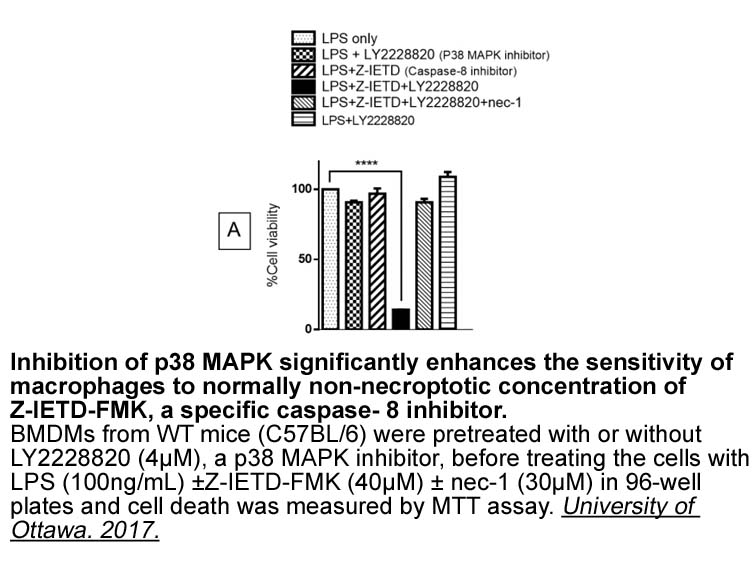
Concluding Remarks Synthetic cytokine biology has become an important research area with novel solutions and ideas for therapeutic approaches, for example, synthekines, fusokines, immunocytokines, neoleukins, MESA receptors, or synthetic Notch or cytokine receptors. In addition to their huge impa
-
In order to gain further insights into the
2020-09-29
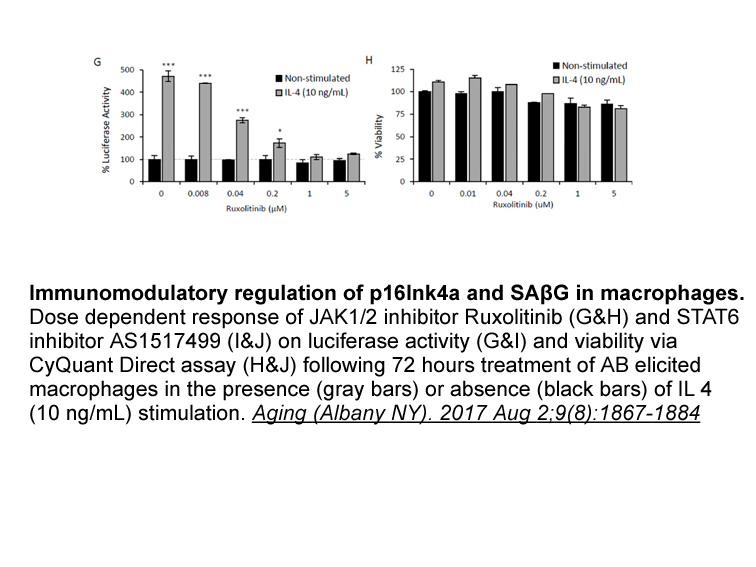
In order to gain further insights into the role of peptidases in B. xylophilus, four cysteine proteases highly secreted by B. xylophilus (Cardoso et al., 2016) were selected four further characterisation. Materials and methods Results and discussion Structural prediction and analysis The i
-
First we synthesized inhibitor following the patent procedur
2020-09-29
First, we synthesized inhibitor following the patent procedure reported by Shewchuk et al. to provide its first complete characterization (see ). Two different synthetic approaches were used to obtain compounds –. The first method relied on the alkylation of the common 5-phenol-2,4-diaminopyrimidine
15899 records 788/1060 page Previous Next First page 上5页 786787788789790 下5页 Last page